The last 12 years here at OIST have been a great journey. From Assistant Professor to Professor and 2 years as Dean, I have “grown up” here as a scientist and manager. Our research has thrived in this unique interdisciplinary environment, and I gave an overview of some of our key research lines in a recent Provost’s lecture. I am most grateful for the many great students, staff, and visitors have spent time in the lab (on the order of a hundred).
With this said, it is time for the lab to enter a new phase. As of September 2024, I (Evan) have moved back to the US to take a position as Professor and Department Chair of Entomology at the University of Maryland. I will be setting up a new biodiversity lab at UMD. The University is located in the suburbs of Washington, D.C., nearby to major institutions such as the Smithsonian Museum of Natural History, the National Institutes of Health, USDA, and many more.
The unit at OIST will continue, and I will lead it as an Adjunct Professor at OIST. I look forward to exciting new opportunities for bi-national collaboration on biodiversity research and education, and we will be working on developing initiatives between the two labs and institutions in the coming months.
If you are interested in working in our group in Maryland or Japan, please reach out.
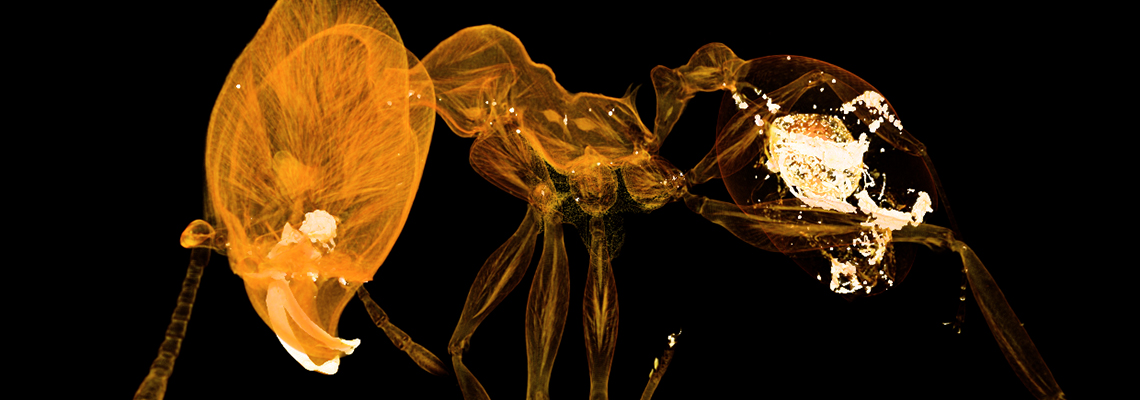
A long-term research goal of our lab is to build a comprehensive database and map of how ants are distributed around the world. We recently published a milestone in this effort, the first high-resolution mapping of richness and rarity. In this study, we compared ant patterns to vertebrates and used machine learning to predict hotspots of undiscovered diversity, which should help guide future research.
One of the things I like about my PhD work is that I get to do a little bit of everything: fieldwork, molecular lab work, and bioinformatics. This helps keep monotony at bay and supplies a variety of interesting problems to solve. Although I like switching back and forth between these tasks, I probably enjoy fieldwork the most. Unfortunately, due to the pandemic, I haven’t had much of an opportunity to conduct fieldwork outside of Okinawa. Now that things are getting better pandemic-wise, I jumped on the opportunity to collect ants in mainland Japan.
Meeting the myrmecology legends in Japan The week before last, lab technician Fumika Azuma and I embarked on a small field expedition to Manazuru forests in Kanagawa Prefecture, just south of Tokyo. My main goal was to collect some Strumigenys hexamera colonies, an Asian species without the iconic trap-jaw mandibles, but rather having the ancestral “gripping” mandibles. It’s known to reproduce parthenogenetically, which is why I wish to establish its colonies in the lab for our Strumigenys comparative genomics work. I was also hoping to collect some species that would serve as outgroups for this project, especially those not found in Okinawa.
Although S. hexamera occurs in Okinawa, I haven’t had luck so far in collecting this rather cryptic, soil-dwelling species. I thus reached out to Dr Keiichi Masuko, who I knew had collected it in the past, to seek advice for where I could collect some colonies. He generously offered to help us and tremendously facilitated the trip. Keiichi, a senior professor at the Senshu University, has been studying Strumigenys ants since before I was born! I was thus delighted to have an opportunity to discuss Strumigenys biology with him; over the years, he has made tons of interesting ecological and natural history observations on this fascinating group of ants, among others. Although Keiichi couldn’t join us in the field, he graciously requested Dr Mamoru Terayama to accompany us, who in turn obliged. Mamoru is a now-retired professor, who has vast experience in the systematics and taxonomy of Japanese ants and wasps. I was thrilled to be collecting ants with and visiting such giants in the Japanese myrmecology community.
Strumigenys hexamera, our target species for this trip. (Photo by Mari, https://kinomari-formica.amebaownd.com)
Fieldwork at Manazuru forests in Kanagawa Boarding the blazing-fast Shinkansen train at Tokyo, we headed to Cape Manazuru, a 30-min drive south of the small city of Odawara in central Japan. Along the way to the cape is a forest patch of moderate size with trails built for adventurous citizens. Although the trails are rather short and not difficult to walk, the forest supports several species of evergreen trees that are quite huge in size. “The trees are at least 300 years old”, I was told by Keiichi. The difference in forests between mainland Japan and Okinawa was stark—the latter, although being more diverse, typically lacks such huge old growth trees because of widespread deforestation and frequent typhoons. The broadleaf evergreen trees in Manazuru forests are primarily camphor, black pine, Itaaji chinkapin, and cane.
Fumika Azuma (left) and I with Mamoru Terayama, at the end of our first day in the field.
The soil arthropod community of the Manazuru forest is rich and abundant. As soon as we began searching, we found several colonies of Carebara, Nylanderia, and Hypoponera, the usual suspects in the ant world. Keiichi had told me that S hexamera nests ~10-20cm deep in the soil. It creates tunnels inside the soil and feeds on the abundant soil arthropods of the forest. Although I had never dug this deep before while looking for Strumigenys, and had mainly been focusing on taking apart rotten logs and flipping rocks, I quickly realized what kind of soil Keiichi was talking about. After digging through 10-15cm of loose soil, I began to find large soil lumps that had clay-like consistency. This type of soil seems ideal for the ants to make tunnels and create compartments to house their brood. Sure enough, within the first hour of beginning fieldwork, I found my first Strumigenys colony. Because Strumigenys species are rather tricky to identify in the field if you haven’t collected them before, I showed it to Mamoru, who immediately confirmed its identity as S hexamera. How exciting! Another highlight of the day included a Discothyrea sauteri colony, which unfortunately only had a queen and one worker, so raising this colony is probably going to be a challenge, not to mention their peculiar diet of arthropod eggs.
The next day we continued our efforts to find more Strumigenys colonies. We succeeded in collecting 5 more colonies, which included S canina and S incerta, neither of which I had collected before. We collected a bunch of other colonies, including a Myrmecina nipponica and Cryptopone sauteri, which might prove useful outgroups. We also collected a worker of Stigmatomma silvestrii, but unfortunately couldn’t locate the colony.
Meeting the members of the Myrmecological Society of Japan Besides meeting Keiichi and Mamoru, we also had the great fortune of having dinner with Dr M Kondoh, the 87-year-old co-founder and current president of the Myrmecological Society of Japan. Dr Yamaguchi, a member of the Society, and his wife, both residing in Odawara, also joined us for the dinner.
Back row: Keiichi Masuko, M Kondoh, me, Fumika. Front row: Mr & Mrs Yamaguchi and Mamoru Terayama. Odawara’s famous Kamaboko fish cake on the right.
It was wonderful to get plugged into the Japanese myrmecologists community and hear stories about their work. The izakaya served a menagerie of delicious food, complete with Kamaboko fish cake, for which Odawara is famous in Japan. All in all, we had a great and productive time in the mainland. Besides these wonderful memories, the M6.1 earthquake I experienced in Tokyo will remain unforgettable. This was the most intense earthquake I have ever experienced and being on the 10th floor didn’t help either. Thankfully there was no tsunami or any damage from the earthquake, but those few seconds were unarguably the scariest of my life. Now back to the lab sorting those ant colonies…
We collected 6 Strumigenys colonies in total and managed to collect one colony of S hexamera. Here they are in their artificial lab nest.
I am on sabbatical this year at Harvard University, where I was lucky enough be named the Mary I. Bunting Fellow at the Radcliffe Institute of Advanced Study. The fellowship program includes 50 scholars to reside here for a year and work on projects in an interdisciplinary environment. The fellowship program has a heavy weight toward Humanities and Social Sciences, which is a great change of pace after being at Natural Science-only OIST for the past years. But luckily they let a few biologists in too (I think every day, at least one must have been by mistake). It’s been fantastic so far to spend time with this amazing group of people at Radcliffe, not to mention some great colleagues and collections at the Museum of Comparative Zoology.
In front of our building, Byerly Hall, which hosts the fellowship program. My office window is under the clock.
Mary I. Bunting, for whom my fellowship is named, was an incredible person. She was a successful microbiologist in a time when when women faced innumerable struggles in being professional scientists. She is best known, however, for her time as president of Radcliffe College when it was a women’s college (versus the all-male Harvard). She was instrumental in highlighting how society had a “climate of unexpectation” for girls and women and contributing to the progress we made toward greater gender equality in the decades since. Both Mary Bunting and Radcliffe have also played important roles in promoting women in academia over the years. In my time at OIST, on the GEDI (Gender Equality, Diversity, and Inclusion) committee, and as Associate Ombudsperson, I have learned a lot about gender and diversity issues that arise in academia, and even more so how those can be amplified in multicultural environments such as OIST. This are better than they were in Mary Bunting’s day, but there is still a long way to go. After 20ish years in academia, I still often notice problems and barriers that contribute to a lack of diversity our fields, many that I had not seen even though they were in plain sight (“A way of seeing is also a way of not seeing”-Kenneth Burke). During my fellowship I will be not only pursuing my “normal” biology research, I will be studying, learning, and thinking about how we can continue to make progress toward a more diverse and inclusive academia. I will bring what I learn back to our lab, OIST, and beyond.
My fellowship is named after Mary I. Bunting, scientist and former president of Radcliffe College, and all around amazing person (image: wikipedia).