Our new paper From species sorting to mass effects: spatial network structure mediates the shift between metacommunity archetypes published in Ecography explores the role of landscape structure in driving biodiversity. Landscape structure becomes important when organisms of interest have ability to move from one place to another, from insects, plants, and animals on terrestrial habitats to marine organisms.
How species are distributed in landscape – how many species inhabit where – and what processes determine the distribution is a big question. Especially, when a large space is considered, this question becomes more complicated, because we need to consider a wider variety of environment and how species distribution differs between places, in addition to the total number of species coexisting, both of which further affect the processes of generating and maintaining biodiversity.
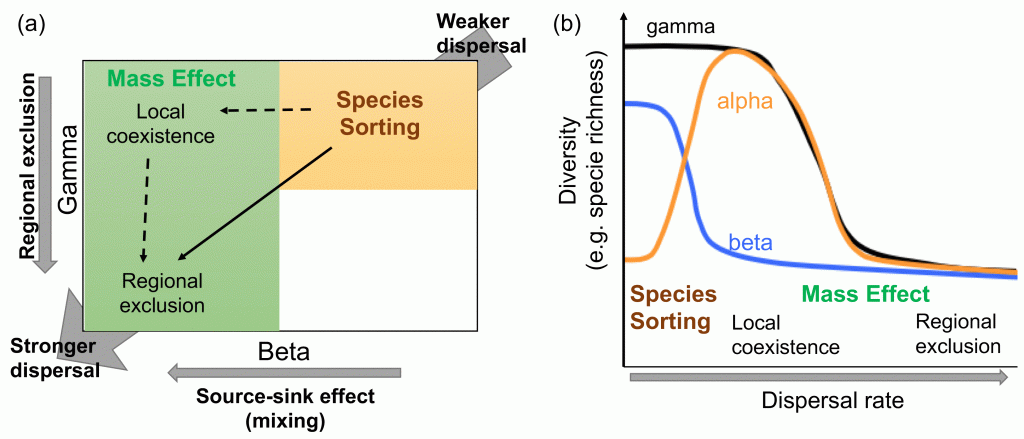
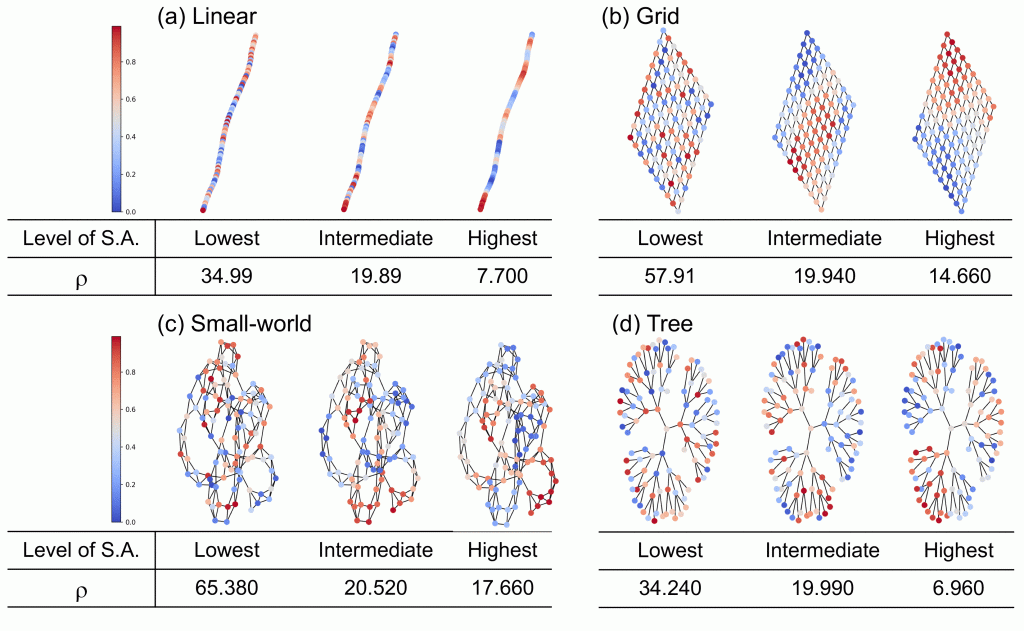
It sounds reasonable to assume simple and regular landscape structures as the first step of exploring the role of spatial structure in ecosystems, expecting that such assumptions would output biodiversity patterns that is average across many different landscape structures. Thus, many theoretical studies addressed this question based on the simple and regular landscape assumption. However, what we found is that such simple and regular assumption generates rather extreme outputs. Check out the paper and find out more details about how this new theory bridges different metacommunity archetypes (species-sorting and mass-effect) and how landscape structure affects the transition between the two archetypes, which then determines what biodiversity patterns emerge.
Yuka Suzuki, PhD Student


 Out in
Out in